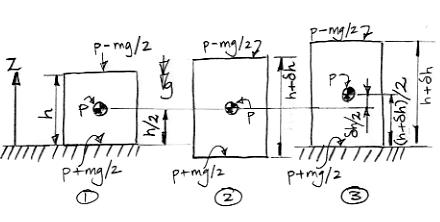
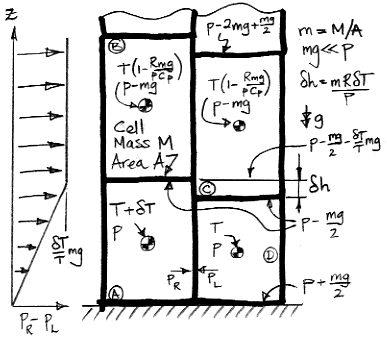
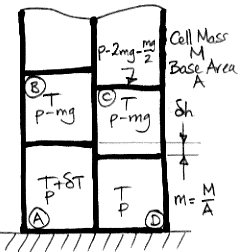
The figure below shows our Circulating Cells simulation program, Version 6.0, which you can download by clicking CC6. With the Left_Only box checked and Planetary Greenhouse heating, the surface cells on the left side receive twice the normal heat from the sun, while those on the right side receive none at all.
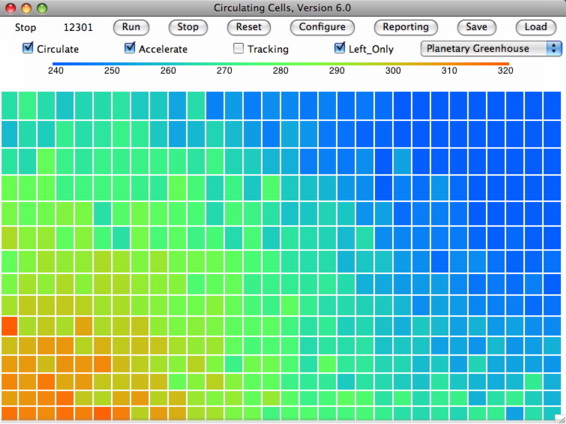
We started the simulation by loading the equilibrium state of the array with both sides receiving heat, which we have saved in PGH_Q001_M00. Following our recent discussion of enthalpy, we recognize this symmetric equilibrium state as the one in which all cells have the same enthalpy. Those at the top have more gravitational potential, but less internal heat and pressure energy, so that the sum of all three forms of energy is the same for all cells, or almost the same.
We checked the Left_Only box and increased Q_heating 0.01 K/s. You may point out that the unit of Q_heating should be Kelvin per iteration, not Kelvin per second, but we recall that one iteration corresponds to one second, so the two are equivalent. With Q_heating at 0.01 K/s, the left-side surface cells warm at 0.02 K/s and those on the right do not warm at all.
We ran the simulation for a million iterations and saved the cell array in PGH_Left_Only. The figure above shows the saved state of the cell array, after another ten thousand iterations. The temperature profile is consistent with a large-area circulation of air powered by heating on the left surface. The heated air rises to the tropopause and moves over to the right side as it radiates its heat into space. Once it has cooled, it descends to the right surface and moves along to the left.
We marked a few cells by clicking on them, and watched them go around. We invite you to do the same. The cells circulate in a clockwise direction. They rise to the tropopause on the left, but hardly ever rise to the tropopause on the right. Nevertheless, we don't see individual cells moving steadily in a clockwise direction across the width and height of the array. Often, cells rise on the extreme left and descend upon the center-left. Cells on the right rise up a little and fall again. They slowly drift to the left, but they do a lot of jumping around along the way.
When averaged over thousands of iterations, the combined movement of the cells is a large clockwise circulation, with a net movement of cells from right to left along the surface. But a simulated person standing on the center surface would not feel a steady breeze blowing from the right side. He would instead feel the wind changing every minute or two, and only by looking at the average wind speed would he be able to conclude that the net movement of air was on-shore.
Our simulation assumes that all momentum generated by circulation is dissipated as viscous heat at the end of each circulation. Thus each circulation affects only the temperature of the cells. No cell can build up momentum that encourages further circulation in the same direction.
We conclude that momentum is one of the driving forces behind the on-shore breezes. Buoyancy alone is not sufficient. If we want our simulation to produce steady winds, we must allow circulating cells to retain some of the momentum they gain during circulation, and we must allow this momentum to influence future circulations of the same cell.